6 - Variations autour des sept domaines transmembranaires :
L'élucidation de la structure moléculaire des RCPGs a montré l'existence de variations autour du thème des sept domaines hydrophobes, qui est caractéristique de cette famille de récepteurs (voir Figure 4 ci-dessous).
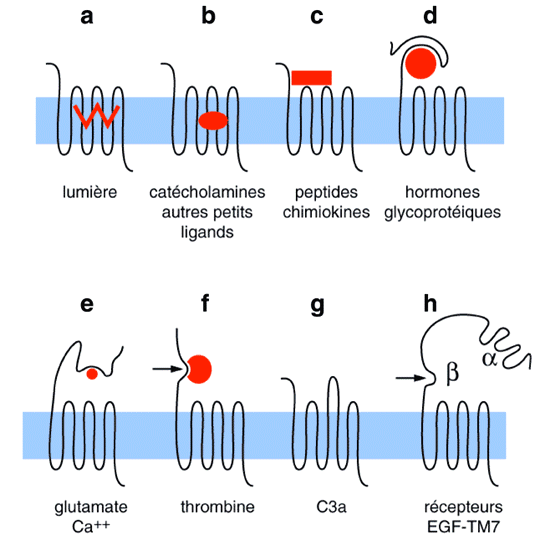
Figure 4 : Variations autour du thème des sept domaines transmembranaires. (D'après Coughlin, 1994 ; Ames et col., 1995 ; McKnight et Gordon, 1996)
Ainsi, les récepteurs visuels (rhodopsine et opsines des cônes) présentent la particularité de posséder un "ligand intrinsèque" (Figure 4a), le chromophore 11-cis-rétinal, qui subit après absorption d'un photon, une photo-isomérisation qui va activer le récepteur (Stryer, 1991).
Les récepteurs des amines et autres petites molécules, ainsi que les récepteurs olfactifs, présentent des régions intra- et extracellulaires peu étendues. Le site de liaison du ligand est souvent localisé à l'intérieur de la poche circonscrite par les sept domaines transmembranaires (Figure 4b).
En revanche, les récepteurs des peptides et des chimiokines lient la plupart du temps leur ligand par l'intermédiaire des régions extracellulaires (Figure 4c).
Les hormones glycoprotéiques (FSH, HCG, LH, TSH), les ions calcium ainsi que l'acide glutamique, reconnaissent la grande extrémité amino-terminale de leurs récepteurs (Figure 4d et 4e).
La thrombine active son récepteur (PARI) par un mécanisme protéolytique (Van Obberghen-Schilling et Pouysségur, 1993). En effet, le clivage spécifique de l'extrémité amino-terminale du récepteur par la thrombine, démasque le vrai ligand, constitué de la nouvelle extrémité du récepteur (Figure 4f et Figure 5). Le clonage ultérieur de deux autres récepteurs activés par des protéases (PAR2 et PAR3) montre une sous-famille en expansion (Coughlin, 1994 ; lshihara et coll., 1997).
Le récepteur de la fraction C3a du complément (Figure 4g) possède une deuxième boucle extracellulaire extrêmement grande (Ames et coll., 1995).
Enfin, un dernier exemple de cette variation concerne les récepteurs de type EGF-TM7 qui comportent une grande région amino-terminale, pouvant être soumise à un clivage protéolytique (Figure 4h) qui conduit à un hétérodimère ab (Mcknight et Gordon, 1996).
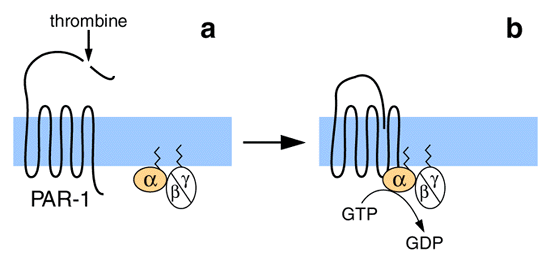
Figure 5 : Activation du récepteur le la thrombine par clivage protéolytique.
Le récepteur de la thrombine (ou Protease-Activated Receptor 1, PAR-1) est clivé au niveau de son extrémité amino-terminale par la thrombine (a). La nouvelle extrémité ainsi libérée constitue le vrai ligand agoniste (b). (D'après Van Obberghen-Schilling et Pouysségur, 1993).